
Synthetic REV-ERB ligands that pharmacologically target the circadian rhythm may be beneficial in the treatment of sleep disorders as well as metabolic diseases.
ABSTRACT
Synchronizing rhythms of behaviour and metabolic processes is important for cardiovascular health and preventing metabolic diseases. The nuclear receptors REV-ERB-α and REV-ERB-β have an integral role in regulating the expression of core clock proteins driving rhythms in activity and metabolism. Here we describe the identification of potent synthetic REV-ERB agonists with in vivo activity. Administration of synthetic REV-ERB ligands alters circadian behaviour and the circadian pattern of core clock gene expression in the hypothalami of mice. The circadian pattern of expression of an array of metabolic genes in the liver, skeletal muscle and adipose tissue was also altered, resulting in increased energy expenditure. Treatment of diet-induced obese mice with a REV-ERB agonist decreased obesity by reducing fat mass and markedly improving dyslipidaemia and hyperglycaemia. These results indicate that synthetic REV-ERB ligands that pharmacologically target the circadian rhythm may be beneficial in the treatment of sleep disorders as well as metabolic diseases.
<center style="box-sizing: border-box; outline: none;">
</center>
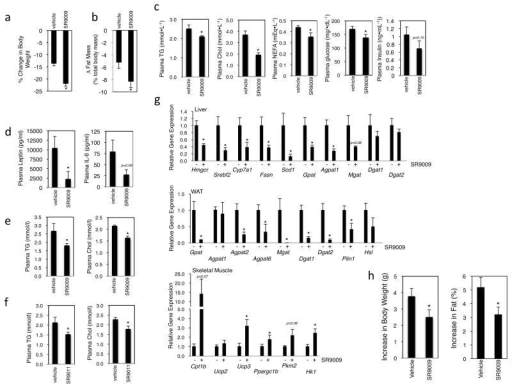
Figure 5: SR9009 treatment results in a decrease in fat mass and in plasma lipids in diet-induced obese micea, Diet-induced obese mice on SR9009 treatment lose weight vs. vehicle treated mice. C57Bl6 mice on a high fat diet were administered SR9009 (100mg/kg, i.p., b.i.d, at CT0 and CT12) for 30 days. b, Diet-induced obese mice on SR9009 treatment exhibit lower fat mass vs. vehicle treated mice. c, Fasting plasma triglycerides (TG), cholesterol (Chol), non-esterified fatty acids (NEFA) and glucose are decreased in SR9009 treated DIO mice. d, Plasma leptin and IL-6 levels from DIO mice e, Fasting plasma TG and Chol in lean C57Bl6 mice. Normal mice were administered 100 mg/kg, i.p., b.i.d. (at CT0 and CT12) SR9009 for 10 days. f, Fasting plasma TG and Chol are decreased by SR9011 treatment in lean C57Bl6 mice. g, Expression of metabolic genes in liver, WAT and skeletal muscle of DIO mice treated with SR9009 as described in a. Gene expression was measured by QPCR and normalized to Cyclophilin b expression. h, SR9009 treatment reduces weigh gain in ob/ob mice. Body weight and body fat content data are shown from ob/ob mice administered SR9009 for 12-days (100 mg/kg, i.p., b.i.d.).* indicates p<0.05. Error bars indicate mean ± s.e.m. and n=6–10 mice
Mentions: Based on the alterations in energy metabolism and gene expression we observed in normal C57BL6 and Balb/c mice, we sought to examine whether a REV-ERBα/β agonist would be efficacious in a rodent model of obesity. We initiated the study with 20-week old C57BL6 mice (average weight = 41g) that had been maintained on a high fat diet for 14 weeks (20% carbohydrate 60% fat). The mice continued on the HF diet and we initiated twice per day dosing (i.p.) of SR9009. While the stress of handling and twice-daily injections caused weight loss in vehicle-treated controls, weight loss of SR9009-treated animals was 60% greater (Fig. 5a). During the treatment period, there was no significant difference in the food intake of SR9009 and vehicle treated animals, although handling itself reduced food intake explaining the weight loss observed in the controls. SR9009 treated mice exhibited a more severe reduction in adiposity (Fig. 5b). In addition to the decrease in fat mass we also observed a 12% decrease in plasma triglycerides (TGs) and a 47% decrease in plasma total cholesterol (Chol) (Fig. 5c). Plasma non-esterified fatty acids (NEFA) were also reduced (23%) along with plasma glucose (19%) in the SR9009 treated animals (Fig. 5c). There was also a trend toward a decrease in plasma insulin levels (35%). Consistent with the decrease in adipocity we also noted an 80% decrease in plasma leptin and a decrease (72%) in the proinflammatory cytokine IL-6 (Fig. 5d). Examination of plasma triglycerides and total cholesterol in lean mice also demonstrated the ability of SR9009 and SR9011 to reduce the levels of these lipids (Figs. 5e, 5f). Consistent with the decreased plasma TGs and total Chol we observed a significant decrease in the expression of genes encoding lipogenic enzymes (Fasn and Scd1) as well as cholesterologenic regulatory proteins (Hmgcr and Srebf2) with SR9009 treatment (Fig. 5g). In the WAT, SR9009 treatment resulted in a decrease in expression of genes encoding enzymes involved in TG synthesis (Fig. 5g) as was also observed in lean mice (Fig. 4d). Similar to our observations in lean mice (Fig. 4c), we observed that the REV-ERB agonist induced the expression of genes involved in fatty acid and glucose oxidation (Cpt1b, Ucp3, Ppargc1b, Pkm2 and Hk1) (Fig. 5g). Taken together with the results from the CLAMS experiments (Fig. 3), these data suggest that REV-ERB agonists increase energy expenditure by increasing fatty acid and glucose oxidation in the skeletal muscle. The gene expression data is also consistent with decreased TG synthesis in the liver and WAT as well as a reduction in hepatic cholesterol synthesis. We also examined the effects of SR9009 in a genetic model of obesity (ob/ob mice) and after 12-days of dosing we observed that SR9009 suppressed the degree of weight gain normally observed in this leptin deficient mouse with no significant alterations in glucose or insulin tolerance (Fig. 5h and data not shown).
ABSTRACT
Synchronizing rhythms of behaviour and metabolic processes is important for cardiovascular health and preventing metabolic diseases. The nuclear receptors REV-ERB-α and REV-ERB-β have an integral role in regulating the expression of core clock proteins driving rhythms in activity and metabolism. Here we describe the identification of potent synthetic REV-ERB agonists with in vivo activity. Administration of synthetic REV-ERB ligands alters circadian behaviour and the circadian pattern of core clock gene expression in the hypothalami of mice. The circadian pattern of expression of an array of metabolic genes in the liver, skeletal muscle and adipose tissue was also altered, resulting in increased energy expenditure. Treatment of diet-induced obese mice with a REV-ERB agonist decreased obesity by reducing fat mass and markedly improving dyslipidaemia and hyperglycaemia. These results indicate that synthetic REV-ERB ligands that pharmacologically target the circadian rhythm may be beneficial in the treatment of sleep disorders as well as metabolic diseases.
<center style="box-sizing: border-box; outline: none;">
</center>
Figure 5: SR9009 treatment results in a decrease in fat mass and in plasma lipids in diet-induced obese micea, Diet-induced obese mice on SR9009 treatment lose weight vs. vehicle treated mice. C57Bl6 mice on a high fat diet were administered SR9009 (100mg/kg, i.p., b.i.d, at CT0 and CT12) for 30 days. b, Diet-induced obese mice on SR9009 treatment exhibit lower fat mass vs. vehicle treated mice. c, Fasting plasma triglycerides (TG), cholesterol (Chol), non-esterified fatty acids (NEFA) and glucose are decreased in SR9009 treated DIO mice. d, Plasma leptin and IL-6 levels from DIO mice e, Fasting plasma TG and Chol in lean C57Bl6 mice. Normal mice were administered 100 mg/kg, i.p., b.i.d. (at CT0 and CT12) SR9009 for 10 days. f, Fasting plasma TG and Chol are decreased by SR9011 treatment in lean C57Bl6 mice. g, Expression of metabolic genes in liver, WAT and skeletal muscle of DIO mice treated with SR9009 as described in a. Gene expression was measured by QPCR and normalized to Cyclophilin b expression. h, SR9009 treatment reduces weigh gain in ob/ob mice. Body weight and body fat content data are shown from ob/ob mice administered SR9009 for 12-days (100 mg/kg, i.p., b.i.d.).* indicates p<0.05. Error bars indicate mean ± s.e.m. and n=6–10 mice
Mentions: Based on the alterations in energy metabolism and gene expression we observed in normal C57BL6 and Balb/c mice, we sought to examine whether a REV-ERBα/β agonist would be efficacious in a rodent model of obesity. We initiated the study with 20-week old C57BL6 mice (average weight = 41g) that had been maintained on a high fat diet for 14 weeks (20% carbohydrate 60% fat). The mice continued on the HF diet and we initiated twice per day dosing (i.p.) of SR9009. While the stress of handling and twice-daily injections caused weight loss in vehicle-treated controls, weight loss of SR9009-treated animals was 60% greater (Fig. 5a). During the treatment period, there was no significant difference in the food intake of SR9009 and vehicle treated animals, although handling itself reduced food intake explaining the weight loss observed in the controls. SR9009 treated mice exhibited a more severe reduction in adiposity (Fig. 5b). In addition to the decrease in fat mass we also observed a 12% decrease in plasma triglycerides (TGs) and a 47% decrease in plasma total cholesterol (Chol) (Fig. 5c). Plasma non-esterified fatty acids (NEFA) were also reduced (23%) along with plasma glucose (19%) in the SR9009 treated animals (Fig. 5c). There was also a trend toward a decrease in plasma insulin levels (35%). Consistent with the decrease in adipocity we also noted an 80% decrease in plasma leptin and a decrease (72%) in the proinflammatory cytokine IL-6 (Fig. 5d). Examination of plasma triglycerides and total cholesterol in lean mice also demonstrated the ability of SR9009 and SR9011 to reduce the levels of these lipids (Figs. 5e, 5f). Consistent with the decreased plasma TGs and total Chol we observed a significant decrease in the expression of genes encoding lipogenic enzymes (Fasn and Scd1) as well as cholesterologenic regulatory proteins (Hmgcr and Srebf2) with SR9009 treatment (Fig. 5g). In the WAT, SR9009 treatment resulted in a decrease in expression of genes encoding enzymes involved in TG synthesis (Fig. 5g) as was also observed in lean mice (Fig. 4d). Similar to our observations in lean mice (Fig. 4c), we observed that the REV-ERB agonist induced the expression of genes involved in fatty acid and glucose oxidation (Cpt1b, Ucp3, Ppargc1b, Pkm2 and Hk1) (Fig. 5g). Taken together with the results from the CLAMS experiments (Fig. 3), these data suggest that REV-ERB agonists increase energy expenditure by increasing fatty acid and glucose oxidation in the skeletal muscle. The gene expression data is also consistent with decreased TG synthesis in the liver and WAT as well as a reduction in hepatic cholesterol synthesis. We also examined the effects of SR9009 in a genetic model of obesity (ob/ob mice) and after 12-days of dosing we observed that SR9009 suppressed the degree of weight gain normally observed in this leptin deficient mouse with no significant alterations in glucose or insulin tolerance (Fig. 5h and data not shown).